УДК 615.357.631.577.157
Изучение генопротекторных свойств препарата "Флараксин" в условиях повреждающего действия тетрахлорметана

Журнал: БИОПОЛИМЕРЫ И КЛЕТКА,
1995, Т. 11, №1, стр 70-80.
ISSN 0233-7657.
Авторы: Ю.И.Губский, Е.Л.Левицкий, И.А.Кулик, А.Г.Горюшко, Р.Г.Примак, Л.Г.Саченко, И.Е.Вистунова.
http://www.biopolymers.org.ua
Реферат
Обнаружено наличие генопротекторных свойств у противоопухолевого препарата растительного происхождения "Флараксин" в условиях повреждения ядерного хроматина печени крыс тетрахлорметаном. По крайней мере частично данный эффект обусловлен антиоксидантными свойствами препарата, что выражается в нормализации течения реакций переокисления липидов во фракциях транскрипционно активного и репрессированного хроматина. Некоторая коррекция структурно-функциональных свойств, хроматина, в условиях повреждения тетрахлорметаном, по всей вероятности, реализуется в результате связывания «Флараксина» с функционально неодинаковыми участками хроматина, причем характер взаимодействия различен для данных участков. Если в транскрипционно активной фракции взаимодействие обусловлено связыванием препарата главным образом с белками хроматина, то в репрессированном хроматине - с ДНК. Образование подобных комплексов «Флараксина» с компонентами хроматина не препятствует реализации генетической информации, однако предохраняет хроматин от повреждения свободными радикалами кислорода и тетрахлорметана.
Введение
Одним из ранних проявлений патологического процесса при отравлении животных тетрахлорметаном (ТХМ) является повреждение ядерного хроматина печени [1, 2]. Предположительно, основным молекулярным механизмом подобного повреждения является накопление свободных радикалов в результате модификации реакций перекисного окисления (ПОЛ) хроматин-связанных липидов [1-3]. Препараты оказывающие антиоксидантное действие в условиях данной патологии, имеют выраженный защитный эффект по отношению к ядерному хроматину печени, т.е. обладают генопротекторной активностью [4,5]. В данной работе приведены результаты исследования генопротекторного действия препарата «Флараксин», разрабатываемого в Институте Фармакологии и Токсикологии АМН Украины. Флараксин – это новое противоопухолевое соединение растительного происхождения фенольной природы, при получении которого использованы неорганические добавки, в частности соли. Предклинические и клинические исследования этого препарата показали, что в лечебных дозах у него отсутствует токсический эффект. В отличие от традиционных противоопухолевых химиотерапевтических средств, он не оказывает выраженного побочного действия на жизненно важные органы и ткани, такие как кровь и кроветворение. Это позволяет применять препарат в крайне запущенных случаях злокачественных меланом, при лечении которых наблюдается выраженный терапевтический эффект.
Материалы и методы
В работе использовали крыссамцов линии Вистар 3-месячного возраста (150-200 г). Животных декапитировали в утренние часы под легким эфирным наркозом, учитывая периодичность митотического цикла. Фракции транскрипционно-активного (ТАХ) и репрессированного (РХ) хроматина печени выделяли, как описано ранее [1,2]. Структурно-функциональное состояние фракций, а также интенсивность реакций ПОЛ регистрировали также спектрофотометрически как отношение оптической плотности диеновых конъюгатов D233 (продуктов ПОЛ) и неокисленной формы липида D215 [6]. Для этого в суспензию липосом из фосфатидилхолина (Сфосфатидилхолина=0,2 мг/мл) вводили раствор Флараксина (С=0,06 мг/мл) и после перемешивания на магнитной мешалке в течение 10 мин записывали спектры поглощения. Затем растворы инкубировали при 37°С, отбирая пробы для спектрофотометрического анализа через определенные промежутки времени. ТХМ вводили внутрибрюшинно в дозе 1,75 мл/кг массы тела животного. Длительность интоксикации составляла 2 ч – период, после которого наблюдается пик генотоксических изменений в хроматине [1-3]. Для изучения влияния Флараксина на процессы ПОЛ, а также на стрктурно-функциональную организацию хроматина животным вводили это вещество в физиологическом растворе в хвостовую вену одновременно с ТХМ, как правило, в дозе 4 мг/кг массы тела (в некоторых экспериментах – 40 мг/кг). В исследованиях in vitro Флараксин добавляли в среду инкубации до конечной концентрации 0,2 и 7,5 мг/мл.
В работе использовали коммерческие препараты бычьего сывороточного альбумина (БСА), ДНК фирмы «Serva» (Германия); лецитин-стандарт (10%-й раствор в спирте) Харьковского предприятия по производству бактериальных препаратов. Липосомы из лецитина готовили, как описано в [7], на диспрегаторе УЗДН-2 (22 кГц, 10 мин).
Антирадикальную активность Флараксина исследовали по методике [8], применяя в качестве стабильного радикала трифенилвердазил (ТФВ) в спиртовом растворе (СТФВ=10-4 М) в области полос поглощения 402 и 740 нм. Долю нейтрализованного свободного радикала при добавлении в раствор Флараксина определяли как отношение ΔD/D0, где ΔD – изменение оптической плотности (при 402 или 740 нм), D0 – оптическая плотность исходного раствора ТФВ.
Собственную флюоресценцию препаратов хроматина изучали в диапазоне 300-450 нм, регистрируя интенсивность спектральной полосы при 320 нм. Структурные изменения в хроматине анализировали с помощью флюоресцентных зондов пирена (перекристаллизованного из метанола) и 1-анилинонафталин-8-сульфоната аммония (1,8-АНС) фирмы «Serva» (Германия), а также явлений индуктивно-резонансного переноса энергии (ИРПЭ) с белковых флюорофоров на пирен в соответствии с [9, 10]. При регистрации ИРПЭ флюоресценцию измеряли при длине волны возбуждения 386 нм и Спирена=2∙10-6 М. Состояние поляризации среды определяли с помощью потенциалчувствительного флюоресцентного зонда diS C3-5 [11]. Флуоресцентные измерения осуществляли на спектрофотометре «Hitachi MPF-4» (Япония) в кварцевых кюветах (1 см) при 20°С. Спектры поглощения хроматина изучали в области 210-370 нм с помощью спектрофотометра «Shimadzu MPS-5000» (Япония) при 20°С. Тепловые ффекты взаимодействия Флараксина с фракциями хроматина и модельными системами регистрировали с использованием микрокалориметра «LKB-2107» (Швеция) в режиме смешения при 26°С.
| Показатель ПОЛ | Контроль | Флараксин, мг/кг | |
|---|---|---|---|
| 4 | 40 | ||
| НЗП | 12417,9 | 9049,3* | 13535,1 |
| НЗП, Δ | 6819,6 | 5226,0 | 9520,9* |
| АЗП | 6335,3 | 5429,5 | 4277,3 |
| Неинициированный контроль | 0 | 0 | 195,9* |
Примечание. Здесь и в табл.2 НЗП-НАДФН-зависимое ПОЛ; НЗП, Δ - его состовляющая, зависимая от нагревания; АЗП – аскорбат-зависимое ПОЛ. *р<0,005 (по сравнению с контролем).
Статистическую обработку полученных данных проводили методами непараметрической статистики [12].
Результаты и обсуждение
Учитывая важную роль модификации процессов ПОЛ в механизмах повреждения клетки ТХМ и его метаболитами [1, 2, 9, 13], представляло интерес изучение влияния Флараксина на эти процессы (табл. 1-5). Данный препарата обладает выраженным антиоксидантным эффектом при определении величины ПОЛ в различных модельных системах in vivo (табл. 1, 3, 5) и in vitro (табл. 2, 4).
| Показатель ПОЛ | Контроль | Флараксин, мг/кг | |
|---|---|---|---|
| 0,2 | 7,5 | ||
| НЗП | 6689,4 | 2614,3* | 2800,0* |
| НЗП, Δ | 4628,7 | 132,3* | 716,7* |
| АЗП | 5808,6 | 181,1* | 96,0* |
| Неинициированный контроль | 623,5 | 397,4* | 804,9* |
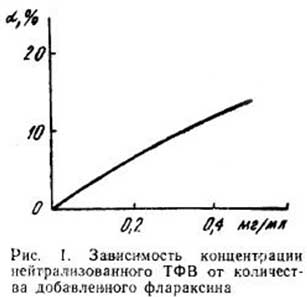
Введение экспериментальным животным Флараксина в дозе 4 мг/кг массы тела приводит к заметному ингибированию НАДФН – индуцированного ПОЛ в гомогенатах печени (табл. 1). Большая доза препарата (40 мг/кг массы тела), наоборот, оказывает прооксидантный эффект на процессы ПОЛ, индуцируемые НАДФН и зависимые от нагревания. При этом увеличивается также интенсивность ПОЛ в неинициированном контроле. При инкубации гомогенатов печени в условиях индукции ПОЛ добавление в среду Флараксина (табл. 2) до конечной концентрации 0,2 и 7,5 мг/мл приводит к достоверному снижению скорости накопления МДА. Эффект выражен в гораздо большей степени, чем in vivo (табл. 1), и четче проявляется в случае меньшей концентрации препарата. При этом обнаружено ингибирование как индуцированного НАДФН, так и аскорбатом ПОЛ, а также липо-переокисления в неинициированном контроле. Отравление животных ТХМ с последующим определением уровня промежуточных продуктов ПОЛ (диеновых конъюгатов) в гомогенатах печени (табл. 3) сопровождается увеличением интенсивности ПОЛ (нейтральных липидов, которые экстрагируются в гептановую фазу липидного экстракта). Вследствие одновременного введения животным Флараксина (4 мг/кг массы тела) наблюдается некоторое снижение уровня ПОЛ как нейтральных липидов, так и фосфолипидов (выделяющих при экстракции в изопропанольную фазу липидного экстракта табл. 3). Количество этих промежуточных продуктов ПОЛ значительно уменьшается при инкубации липосом с Флараксином в условиях стимуляции липопереокисления in vitro (табл. 4). Исследование антирадикальной активности Флараксина показало, что он может служить «ловушкой» стабильного радикала ТФВ (рис. 1). Это полностью соответствует результатам биохимических исследований.
При изучении процессов спонтанного и индуцированного ПО во фракциях РХ и ТАХ в условиях интоксикации животных ТХМ и введения Флараксина было обнаружено следующее (табл. 5). Введение ТХМ вызывает угнетение процессов как спонтанного, так и индуцированного НАДФН ПОЛ во фракции ТАХ. Столь парадоксальный, на первый взгляд, факт может быть связан как с изменением жирнокислотного состава этой фракции хроматина в результате интоксикации [13], так и с изменением ее физико-химических свойств [14], в результате чего, возможно, снижается доступность жирнокислотных остатков хроматин-связанных липидов действию окисляющих (повреждающих) агентов, в качестве которых могут быть свободные радикалы ТХМ и кислорода.
Во фракции низкоактивного хроматина введение ТХМ вызывает стимуляцию ПОЛ, индуцированного внесением в среду инкубации аскорбата. Подобная разнонаправленность изменений интенсивности реакций ПОЛ во фракциях хроматина при однократном введении ТХМ может быть обусловлена различными вариациями жирнокислотного состава и физико-химических свойств этих фракций под влиянием интоксикации [13, 14]. Одновременное с ТХМ введение животным Флараксина оказывает нормализующий эффект на течение процессов ПОЛ в обеих фракциях хроматина. По всей вероятности, антиоксидантный и прооксидантный эффекты Флараксина по отношению к различным фракциям хроматина могут быть реализованы вследствие изменения (нормализации) структурно-функциональных свойств хроматина, нарушенных в результате интоксикации ТХМ. Подробное предположение подтверждается результатами исследований, представленных в табл. 6 и 7 на рис. 2-5. В табл. 6 приведены интегральные биохимические параметры структурной организации РХ и ТАХ в условиях интоксикации ТХМ и введения Флараксина. При введении ТХМ отмечается увеличение доли ТАХ и соответственно снижение РХ, а также увеличение отношения белок/ДНК во фракции ТАХ. Введение животным флараксина нормализует данные параметры структуры хроматина. Подобные изменения структуры фракций хроматина не сопровождаются увеличением их транскрипционной активности в условиях in vitro (табл. 6), что позволяет предположить наличие нарушений структурно-функциональной организации хроматина в результате интоксикации ТХМ.
| Фаза липидного экстракта | Контроль | ТХМ | ТХМ + Флараксин |
|---|---|---|---|
| Гептановая | 55,9 | 122,7* | 113,4* |
| Изопропанольная | 833,0 | 794,3 | 561,8* |
Примечание *р<0,05 (по сравнению с контролем)
| Условия эксперимента | Время инкубации, ч | ||
|---|---|---|---|
| 1 | 24 | 48 | |
| Липосомы | 0,239 | 0,281 | 0,655* |
| Липосомы + флараксин | 0,283 | 0,332 | 0,187* |
Примечание *р<0,05 (по сравнению с1-ч инкубацией)
| Показатель ПОЛ | РХ | ТАХ | ||||
|---|---|---|---|---|---|---|
| Контроль | ТХМ | ТХМ + Флараксин |
Контроль | ТХМ | ТХМ + Флараксин |
|
| НЗП | 889,4 | 129,3 | 1279,5 | 459,5 | 274,6 | 317,0 |
| НЗП, Δ | 0 | 0 | 0 | 14,7 | 1,0* | 10,5 |
| АЗП | 422,1 | 653,0* | 442,3 | 62,0 | 68,2 | 78,4 |
| Неинициированный контроль | 258,8 | 250,9 | 225,0 | 55,9 | 17,8* | 41,6 |
| Диеновые конъюгаты: Гептановая фаза липидного экстракта | 0 | 0 | 0 | 0 | 0 | 0 |
| Диеновые конъюгаты: Изопропанольная фаза липидного экстракта | 146,8 | 132,6 | 159,5 | 272,8 | 146,8* | 248,7 |
Примечание *р<0,05 (по сравнению с контролем)
ТХМ вызывает изменение функциональной активности хроматина, которое выражается в резком угнетении РНК-полимеразной активности в РХ и ТАХ. При этом наблюдается снижение эндогенной активности как фракции ферментов, обогащенной РНК-полимеразой II. Введение Флараксина оказывает частичный нормализующий эффект на функциональную активность хроматина, в особенности репрессированного. Во фракции РХ генопротекторный эффект препарата связан с резкой стимуляцией активности фракции, обогащенной РНК-полимеразой I. Вследствие этого, хотя активность фракции, обогащенной РНК-полимеразой II, и остается сниженной по сравнению с контролем в печени животных, которым вводили ТХМ и Флараксин, тем не менее, тотальная РНК-полимеразная активность достоверно не отличается у них по сравнению с контрольными значениями. Во фракции ТАХ введение Флараксина оказывает незначительный генопротекторный эффект на эндоенную РНК-полимеразную активность хроматина, который проявляется в большей степени по отношению к активности фракции ферментов, обогащенной РНК-полимеразой I.
| Показатель ПОЛ | РХ | ТАХ | ||||
|---|---|---|---|---|---|---|
| Контроль | ТХМ | ТХМ + Флараксин |
Контроль | ТХМ | ТХМ + Флараксин |
|
| Доля фракции, % | 88,4 | 82,5* | 85,7 | 11,7 | 17,5* | 14,3 |
| Отношение белок / ДНК | 1,4 | 1,6 | 1,5 | 11,0 | 13,0* | 12,2 |
| Активность эндогенных РНК-полимераз (расп/мин на 1 мг ДНК): | ||||||
| Тотальная | 489413 | 233288* | 365725 | 2723581 | 949952* | 1055011* |
| РНК-полимераза I | 34333 | 11630* | 11630* | 689796 | 270769* | 327847* |
| РНК-полимераза II | 465080 | 221658* | 246288* | 1997758 | 512693* | 596649* |
Примечание *р<0,05 (по сравнению с контролем)
В целом действие Флараксина на эндогенную РНК-полимеразную активность фракции РХ и ТАХ в условиях интоксикации ТХМ проявляется в гораздо меньшей степени, чем таковое по отношению к процессам ПОЛ. Это позволяет предположить, помимо свободнорадикальных механизмов повреждения хроматина ТХМ, также наличие дополнительных механизмов нарушения функций генома, которые реализуются, например, в результате связывания ТХМ с РХ и ТАХ [2]. Последнее способно вызывать изменение структуры двойной спирали ДНК в хроматине, что, в свою очередь, может искажать процесс транскрипции. Возможно также нарушение процесса считывания генетической информации на этапе трансляции в клетках отравленных животных [15], сопровождающееся синтезом функционально неполноценных молекул РНК-полимераз и, как следствие, искажением синтеза м-РНК и р-РНК. При этом нельзя исключить того, что корригирующий эффект на процессы трансляции у Флараксина отсутствует.
Итак, представленные результаты биохимических исследований свидетельствуют о нарушении функциональной активности фракционированного хроматина печени под влиянием ТХМ. Частично этот эффект обусловлен модификацией свободнорадикальных реакций переокисления хроматин-связанных липидов. Предположительно, интоксикация изменяет таким образом структуру фракций хроматина, что в РХ проявляется индуцирующий эффект аскорбата на процессы ПОЛ, в то время как в ТАХ жирнокислотные остатки хроматин-связанных липидов становятся в результате интоксикации менее доступными прооксидантному действию НАДФН по сравнению с контролем. Искажение структуры хроматина под влиянием действия ТХМ может также являться одним из факторов, обуславливающих изменение его эндогенной РНК-полимеразной активности. Можно предположить, что частичный корригирующий эффект Флараксина на течение процессов ПОЛ в хроматине и на его РНК-синтезирующую активность может быть также обусловлен структурной реорганизацией хроматина под влиянием введения данного препарата. Для проверки изложенных предположений были проведены эксперименты, в которых определяли структурное состояние фракций хроматина в условиях интоксикации животных ТХМ и введения Флараксина с фракциями РХ и ТАХ in vitro физико-химическими методами. Результаты этих экспериментов представлены в табл. 7 и на рис. 2-5.
| Условия эксперимента | Д210/Д260 | Д230/Д260 | Условия эксперимента | Д210/Д260 | Д230/Д260 |
|---|---|---|---|---|---|
| РХ | ТАХ | ||||
| Контроль | 2,54 | 0,78 | Контроль | 3,20 | 2,0 |
| ТХМ | 2,57 | 0,79 | ТХМ | 3,26 | 2,31* |
| ТХМ + флараксин | 2,53 | 0,77 | ТХМ + флараксин | 3,44 | 2,05 |
Примечание *р<0,05 (по сравнению с контролем)
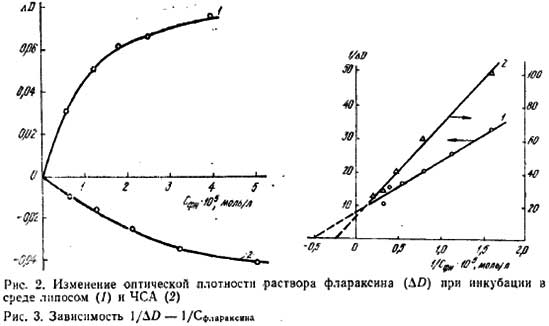
В исследуемой области спектров поглощения хроматина наблюдаются полосы, обусловленные хромофорами молекул ДНК, белков, а также липидов, структура и организация которых при интоксикации могут нарушаться [4, 14]. В табл. 7 приведены спектральные параметры – относительные оптические плотности D/D260 (λ 210 и 230 нм), отражающие изменения в белковых или липидных участках хроматина (отношения белок/липид) при повреждениях хроматина, вызванных интоксикацией ТХМ, а также в условиях введения Флараксина. Из представленных данных следует, что интоксикация ТХМ мало сказывается на спектральных характеристиках фракции РХ, в то время как в спектрах ТАХ наблюдается увеличение D230/D260, свидетельствующее о возможном возрастании содержания липидов (белков) во фракции в условиях интоксикации. Введение животным Флараксина нормализует данный показатель. Отношение D210/D260 не изменяется существенным образом при интоксикации ТХМ и введении Флараксина ни в РХ, ни в ТАХ, что подтверждает неизменность суммарного содержания неокисленных фосфолипидов во фракциях. Методами спектрофотометрии и микрокалориметрии была исследована способность Флараксина к комплексообразованию с РХ и ТАХ. Как показало спектрофотометрическое титрование растворов хроматина (Сбелка=22,8 и 20,7 мкг/мл для ТАХ и РХ соответственно) раствором Флараксина, при весовых соотношениях Флараксина и белка менее 1:1 наблюдается гиперхромный эффект при λ=290 нм; при больших – снижение оптической плотности, что может указывать на межмолекулярное взаимодействие Флараксина с хроматином (данные не приведены). Регистрируемые изменения оптической плотности малы и недостаточны для расчета констант равновесия реакций. Поэтому для выяснения вопроса о месте локализации Флараксина в хроматине было проведено спектрофотометрическое титрование Флараксином модельных систем – лецитиновых липосом (0,25 мг/мл) и человеческого сывороточного альбумина (ЧСА, 0,05 мг/мл).
На рис. 2 приведены зависимости ΔD-Cфлараксина, где ΔD=Di-D0 (Di и D0 - оптическая плотность Флараксина при длине волны 278 и 290 нм без и в присутствии субстрата соответственно). При инкубации Флараксина в среде липосом в его спектре поглощения наблюдается возрастание оптической плотности при 290 нм (кривая I) и снижение при 350 нм, в то время как в растворе, содержащем ЧСА, отмечается снижение оптической плотности в данной области (D278, кривая 2), что может указывать на различные механизмы взаимодействия Флараксина с фосфолипидным бислоем и белковыми молекулами.
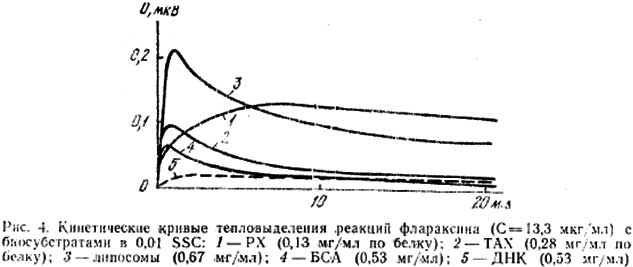
На основании данных спектрофотометрии в соответствии с [16] были построены зависимости 1/ΔD-1/С (рис. 3) для липосом (кривая I) и ЧСА (кривая 2), из которых были оценены константы равновесия их реакции с Флараксином, равные соответственно: (5,0±0,5)∙104 и (2,4±0,5)∙104 М-1∙л. Одинаковый порядок величин Кp свидетельствует об образовании Флараксином прочных комплексов как с ЧСА, так и с липидным компонентом хроматина (на основании его связывания с липосомами), что, вероятно, свидетельствует о наличии конкуренции за места связывания с этими образованиями in vivo. Взаимодействие Флараксина с хроматином и его компонентами подтверждают также микрокалориметрические исследования. На рис. 4 приведены кинетические кривые тепловыделения реакции Флараксина с РХ (кривая 1) и с ТАХ (кривая 2). Как можно видеть, кинетика тепловыделения и интенсивность различны для РХ и ТАХ: в первом случае максимум теплового эффекта наблюдается через 2-3 мин после смешивания растворов, во втором – через 8-10 мин. Для выяснения природы взаимодействия исследованных реагентов эти данные были сопоставлены с результатами, полученными для модельных систем: фосфатидилхолиновых липосом (рис. 4, кривая 3), бычьего сывороточного альбумина (БСА, кривая 4) и ДНК (кривая 5). Для реакции с РХ характер зависимости близок к ДНК-взаимодействию, в то время как во фракции ТАХ кинетические кривые аналогичны таковым для липосом или БСА. Величины теплового эффекта взаимодействия Флараксина с ТАХ и БСА близки, что дает возможность предположить доминирующий вклад в этот процесс белкового компонента хроматина. Такое предположение подтверждается полученными данными при исследовании ИРПЭ в системах хроматин-пирен и хроматин-флараксин-пирен. В соответствии с [9], в присутствии пирена при возбуждении белковой флюоресценции хроматина (286 нм) имеет место ИРПЭ с белковых флюорофоров на акцептор-флюоресцентный зонд пирен. По результатам тушения флюоресценции были рассчитаны вероятности W переноса энергии с белковых флюорофоров РХ и ТАХ на пирен (0,40 и 0,18 соответственно). При инкубации хроматина в присутствии Флараксина (5 мкг/мл) наблюдается еще более эффективное тушение белковой флюоресценции хроматина. Оцененные вероятности переноса энергии в таких системах оказались равными для РХ и ТАХ 0,71 и 0,82 соответственно. Более высокие значения W в присутствии Флараксина обусловлены, по-видимому, пертурбациями в структуре белковых флюорофоров хроматина, сопровождающимися уменьшением расстояния между донором и акцептором энергии. Подобные изменения возможны как в условиях более компактной структуры белков хроматина, модифицированной Флараксином, так и при большей вероятности встраивания пирена в липидную составляющую хроматина при снижении ПОЛ (учитывая антиоксидантное действие Флараксина). Большее значение W в случае ТАХ по сравнению с РХ в присутствии Флараксина согласуется с предположениями, сделанными нами о природе его взаимодействия с РХ и ТАХ на основании данных микрокалориметрии. Было показано, что взаимодействие фларасина с ТАХ происходит преимущественно с участием белковых структур. Не столь значительное возрастание W (менее чем в 2 раза) в случае РХ связано, очевидно, со взаимодействием Флараксина с ДНК в этой фракции хроматина.
Более того, Флараксин может в ИРПЭ выступать в качестве посредника между донором и акцептором ввиду того, что энергетический уровень Флараксина (λфлюоресценции=350 нм) является промежуточным между триптофанилом (λфлюоресценции=330 нм) и пиреном (λфлюоресценции =370 нм и выше).
Таким образом, проведенные исследования свидетельствуют о том, что внедрение Флараксина в структуру хроматина приводит к структурно-динамическим изменениям в глубинных гидрофобных участках хроматина.
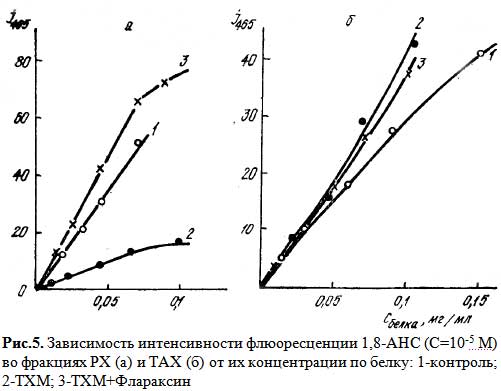
Изменения поверхностных свойств хроматина под влиянием Флараксина изучали с помощью анионного зонда 1,8-АНС. На рис. 5 приведены зависимости интенсивности флюоресценции зонда от концентрации хроматина (по белку) для фракций РХ и ТАХ. В РХ при интоксикации ТХМ значительно снижается способность зонда к встраиванию (кривая 2), что может быть как результатом возрастания отрицательного заряда поверхности липидного компонента хроматина, так и уплотнениям поверхности в результате структурных пертурбаций. Введение животным Флараксина способствует нормализации данного параметра: встраивание АНС происходит более интенсивно (кривая 3), чем в контрольных образцах (кривая 1). В отличие от РХ, в ТАХ интоксикация животных ТХМ приводит к возрастанию встраивания зонда (кривая 2), что может свидетельствовать об уменьшении отрицательного заряда поверхности липидного компонента хроматина либо о ее разрыхлении. Инъекция животным Флараксина несколько снижает этот эффект (кривая 3), приближая значения интенсивности свечения зонда к контролю (кривая 1). Следует учесть, однако, что определенный вклад в рассматриваемые процессы встраивания АНС может вносить различное содержание белка в РХ и ТАХ. По-видимому, взаимодействие Флараксина и ДНК во фракциях РХ в условиях in vivo модифицирует его поверхность, повышает эффект встраивания зонда, в отличие от ТАХ, где количество белков значительно выше. Поэтому образование комплекса белок-флараксин способствует уплотнению поверхности, вследствие чего возможность встраивания зонда уменьшается.
Следовательно, в настоящей работе показано, что Флараксин вызывает изменение свойств как внутренних глубинных (гидрофобных) участков, так и поверхности хроматина благодаря своей способности образовывать межмолекулярные комплексы с белками, липидами, а также с ДНК хроматина. Поскольку РХ и ТАХ отличаются по белковому, липидному составу, а также по содержанию ДНК [17], наблюдаются значительные различия в кинетике их взаимодействия с Флараксином.
Суммируя изложенное выше, можно заключить, что в результате изучения влияния препарата «Флараксин» на структурно-функциональную организацию ядерного хроматина печени крыс в условиях интоксикации ТХМ было обнаружено наличие у этого препарата генопротекторных свойств. По крайней мере частично, данный эффект обусловлен антиоксидантными свойствами препарата, что находит свое выражение в нормализации течения реакций переокисления хроматин связанных липидов. Выявленное в экспериментах на модельных системах связывание препарата с ДНК в РХ и белками и липидами в ТАХ может лежать в основе реализации его генопротекторного эффекта, препятствуя, таким образом, свободно радикальному повреждению компонентов ядерного хроматина либо изменяя конформацию этих компонентов, вследствие чего они становятся невосприимчивыми к действию свободных радикалов, либо связывая свободные радикалы и, значит, предохраняя структуру хроматина от повреждения.
Наличие по отношению к ядерному хроматину защитного эффекта у препарата «Флараксин» коррелирует со снижением смертности животных в результате интоксикации ТХМ (68,9% без Флараксина и 33,3% с препаратом p<0,05).
Ю.І.Губський, Є.Л. Левицький, І.О.Кулик,
Г.Г. Корюшко, Р.Г.Примак, Л.Г.Саченко, І.Є.Вістунова
ВИВЧЕННЯ ГЕНОПРОТЕКТОРНИХ ВЛАСТИВОСТЕЙ ПРЕПАРАТУ «ФЛАРАКСИН» ЗА УМОВ ПОШКОДЖУЮЧОЇ ДІЇ ТЕТРАХЛОРМЕТАНУ
Резюме
Виявлено наявність генопротекторных властивостей у протипухлинного препарату «Флараксин» за умов пошкодження хроматину тетрахлоретаном. Щонайменше частково цей ефект зумовлений антиоксидантними властивостями препарату, що виражається у нормалізації реакцій переокислення ліпідів у фракціях транскрипційно активного та репресованого хроматину. Часткова корекція структурно-функціональних властивостей хроматину за умов ушкодження тетрахлоретаном найімовірніше реалізується внаслідок зв’язування флараксину з функціонально неоднаковими ділянками хроматину, причому характер взаємодії відрізняється для різних ділянок. Якщо у транскрипційно активній фракції взаємодія обумовлена зв’язуванням препарату головним чином з білками хроматину, то у репресованому хроматині – з ДНК. Утворення подібних комплексів флараксину з компонентами хроматину не перешкоджає реалізації генетичної інформації, однак запобігає ушкодженню хроматину вільними радикалами кисню та тетрахлоретану.
Yu.I.Gubskiy, E.L.Levitsky, I.A.Kulick,
A.G.Goryushko, R.G.Primak, L.G.Sachenko, I.E.Vistunova
STUDY OF GENOPROTECTIVE FATURES OF PREPARATION «FLARAKSIN» IN CONDITIONS OF TETRACHLOROMETHANE DAMAGE ACTION
Summury
The presence of genoprotective features of antitumor preparation “Flaraksin” in conditions of the rat liver nuclear chromatin damage by tetrachlormethane was revealed. At least partially this effect may be conditioned by antioxidant action of the preparation. This action is expressed as a normalization of the lipoperoxidation reactions in the transcriptionally active and repressed chromatin fractions. Partial correction of chromatin structural and functional characteristics in condition of chromatin damage by tetrachloromethane is realized probably in result of flaraksin bounding to functionally different chromatin sites. The character of this bounding is various in these sites. If in active fraction interaction is caused mainly by chromatin proteins, then in repressed one – DNA. Such chromatin-flaraksin complexes do not prevent the genetic information realization, however they protect the nuclear chromatin from damage by oxygen and tetrachloromethane free radicals.
СПИСОК ЛИТЕАТУРЫ
- Левицкий Е.Л., Фальковская Е.Н. Ранние изменения в хроматине печени при интоксикации тетрахлорметаном // Фармакология и токсикология. Респ. межведомств. Сборник. – Киев : Здоровья, 1992, - С. 32-35.
- Губский Ю.И., Левицкий Е.Л., Жила В.А. и др. Молекулярные механизмы повреждения фракционированного хроматина печени тетрахлорметаном // Вопр. мед. химии. – 1992.-38, №3.-С. 54-58.
- Губский Ю.И., Левицкий Е.Л. Механизмы перекисного окисления липидов фракций хроматина печени крыс // Биополимеры и клетка.- 1993.-9, №5.-С. 34-43.
- Губский Ю.И., Левицкий Е.Л., Примак Р.Г.и др. Влияние витамина Е на структурно-функциональную организацию хроматина печени в условиях повреждения тетрахлорметаном // Биополимеры и клетка.-1993-9, №3.-С. 27-34.
- Губский Ю.И., Левицкий Е.Л., Холодова Ю.Д. и др. Механизмы генопротекторного действия препарата на основе фитоэкдистероидов (БТК-8Л) в условиях повреждения хроматина тетрахлорметаном // Укр. биохим. Журн.-1993.-65, №6.-С. 75-83.
- Владимиров Ю.А., Арчаков А.И. Перекисное окисление липидов в биологических мембранах.-М.: Наука, 1972.-252 с
- Лебедь О.И., Стефанов А.В., Примак Р.Г. Влияние условий ультразвуковой обработки на характеристики формирующихся липосом // Укр. биохим. журн.-1989.-61, №3.-С. 96-101.
- Мельничук В.А., Починок Т.В., Портнягина В.А. и др. Антиокислительная активность некоторых лекарственных средств растительного происхождения // Фармакология и токсикология. Респ. межведомст. Сборник – Киев:Здоровья, 1988.-С. 79-83.
- Владимиров Ю.А., Добрецов Г.Е. Флюоресцентные зонды в исследовании клеток, мембран и липопротеинов. - М.: Медицина, 1980.-320 с.
- Владимиров Ю.А., Добрецов Г.Е. Флюоресцентные зонды в исследовании клеток, мембран и липопротеинов. - М.: Наука, 1989.-277 с.
- Постнов Ю.В., Орлов С.Н. Первичная гипертензия как патология клеточных мембран. – М.: Медицина, 1987. – 192 с.
- Ашмарин И.П., Васильев Н.Н., Амбросов В.А. Быстрые методы статистической обработки и планирование экспериментов. – Л.: Изд-во ЛГУ, 1975. – 78 с.
- Губский Ю.И., Левицкий Е.Л., Волков Г.Л. Жирнокислотный состав фракций хроматина печени крыс в условиях стимуляции перекисного окисления липидов // Укр. биохим. журн. – 1991.-63, №1.- С. 87-91.
- Губский Ю.И., Левицкий Е.Л., Примак Р.Г.. Изменение структурного состояния фракционированного хроматина печени при активации перекисного окисления липидов // Биополимеры и клетка. – 1991.-7, №3.- С. 89-94.
- Губский Ю.И. Коррекция химического поражения печени. – Киев : Здоров’я, 1989.- 168 с.
- Свердлова О.В. Электронные спектры в органической химии.-Л.: Химия, 1985.-248 с.
- Левицкий Е.Л., Губский Ю.И., Чабанный В.Н. и др. Биохимическая характеристика фракций транскрипционно активного и репрессированного хроматина печени крыс // Биополимеры и клетка.-1993.-9, №6.-С. 11-21.
Ин-т фармакологии и токсикологии АМН Украины, Киев. Получено 14.06.94
